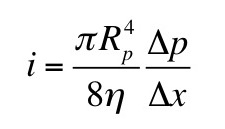|
| Mathematical Physiology, by James Keener and James Sneyd. |
It can be argued that of all the biological sciences, physiology is the one in which mathematics has played the greatest role. From the work of Helmholtz and Frank in the last century through to that of Hodgkin, Huxley, and many others in this century [the first edition of MP was published in 1998], physiologists have repeatedly used mathematical methods and models to help their understanding of physiological processes. It might thus be expected that a close connection between applied mathematics and physiology would have developed naturally, but unfortunately, until recently, such has not been the case.If you substitute the words “physics” for “mathematics,” “physical” for “mathematical,” and “physicist” for “mathematician,” you would almost think that this preface had been written by Russ Hobbie for an early edition of IPMB.
There are always barriers to communication between disciplines. Despite the quantitative nature of their subject, many physiologists seek only verbal descriptions, naming and learning the functions of an incredibly complicated array of components; often the complexity of the problem appears to preclude a mathematical description. Others want to become physicians, and so have little time for mathematics other than to learn about drug dosages, office accounting practices, and malpractice liability. Still others choose to study physiology precisely because thereby they hope not to study more mathematics, and that in itself is a significant benefit. On the other hand, many applied mathematicians are concerned with theoretical results, proving theorems and such, and prefer not to pay attention to real data or the applications of their results. Others hesitate to jump into a new discipline, with all its required background reading and its own history of modeling that must be learned.
But times are changing, and it is rapidly becoming apparent that applied mathematics and physiology have a great deal to offer one another. It is our view that teaching physiology without a mathematical description of the underlying dynamical processes is like teaching planetary motion to physicists without mentioning or using Kepler’s laws; you can observe that there is a full moon every 28 days, but without Kepler’s laws you cannot determine when the next total lunar or solar eclipse will be nor when Halley’s comet will return. Your head will be full of interesting and important facts, but it is difficult to organize those facts unless they are given a quantitative description. Similarly, if applied mathematicians were to ignore physiology, they would be losing the opportunity to study an extremely rich and interesting field of science.
To explain the goals of this book, it is most convenient to begin by emphasizing what this book is not; it is not a physiology book, and neither is it a mathematics book. Any reader who is seriously interested in learning physiology would be well advised to consult an introductory physiology book such as Guyton and Hall (1996) or Berne and Levy (1993), as, indeed, we ourselves have done many times. We give only a brief background for each physiological problem we discuss, certainly not enough to satisfy a real physiologist. Neither is this a book for learning mathematics. Of course, a great deal of mathematics is used throughout, but any reader who is not already familiar with the basic techniques would again be well advised to learn the material elsewhere.
Instead, this book describes work that lies on the border between mathematics and physiology; it describes ways in which mathematics may be used to give insight into physiological questions, and how physiological questions can, in turn, lead to new mathematical problems. In this sense, it is truly an interdisciplinary text, which, we hope, will be appreciated by physiologists interested in theoretical approaches to their subject as well as by mathematicians interested in learning new areas of application.
Many of the topics in MP overlap those in IPMB: diffusion, bioelectricity, osmosis, ion channels, blood flow, and the heart. MP covers additional topics not in IPMB, such as biochemical reactions, calcium dynamics, bursting pancreatic beta cells, and the regulation of gene expression. What IPMB has that MP doesn’t is clinical medical physics: ultrasound, x-rays, tomography, nuclear medicine, and MRI. Both books assume a knowledge of calculus, both average many equations per page, and both have generous collections of homework problems.
Which book should you use? Mathematical Physiology won an award, but Intermediate Physics for Medicine and Biology has an award-winning blog. I’ll take the book with the blog. I bet I know what Frankie will say: “I’ll take both!”