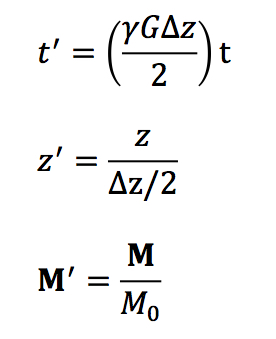During the visit, I noticed how physics underlies most of Haines’s work in the clinic. Much of this physics is described in Intermediate Physics for Medicine and Biology. Russ Hobbie and I discuss the electrical behavior of the heart and the electrocardiogram in Chapter 7, arrhythmias such as fibrillation in Chapter 10, and the bioheat equation governing the tissue temperature in Chapter 14.
 |
| Cardiac Electrophysiology: From Cell to Bedside, 4th Ed., Edited by Zipes and Jalife. |
The rationale of ablation is that, for every arrhythmia, there is a critical region of abnormal impulse generation or propagation that is required for that arrhythmia to be sustained clinically. If that substrate is irreversibly altered or destroyed, then the arrhythmia should not occur spontaneously or with provocation. To accomplish this with a catheter, several criteria must be met. The technology needs to be controllable: Big enough to incorporate the target but small enough to minimize collateral damage. It needs to be affordable and adaptable to equipment conventionally found in the electrophysiology (EP) suite. Despite considerable experience and experimentation with a variety of catheter ablation technologies, ablation with radiofrequency (RF) electrical energy emerged and has persisted as the favored modality. The study of the mechanisms of RF energy heating and the tissue’s response to this injury will give insight into these and other phenomena and should allow the operator to optimize procedure outcome.Let me describe some of the physics of catheter ablation.
- The Catheter. A catheter is used to place the lead used for ablation into the heart. Usually it’s inserted into a vein in the leg, and then snaked through the vessels into the right atrium. (Ablating tissue in the left atrium is trickier; you may have to create a small hole between the atria by doing a transseptal puncture.) Catheterization is less invasive than open heart surgery, so some patients can avoid even a single night in the hospital after treatment.
- Radiofrequency Energy. Ablation is performed using electrical energy with a frequency between 0.3 and 1 MHz (in the frequency band of AM radio). These frequencies are too high to cause direct electrical stimulation of muscles or nerves. The mechanism of ablation is Joule heating, like in your toaster, which raises the temperature of the tissue within a few millimeters of the lead tip.
- Lesion Formation. Cells become irreversibly damaged at temperatures on the order of 50° C. The temperature of the lead tip is kept below 100° C to avoid boiling the plasma and coagulating proteins.
- Atrial Fibrillation. Atrial fibrillation is the most common arrhythmia treated with ablation. Fibrillation means that the electrical wave fronts propagate in a irregular and chaotic way, so the mechanical contraction is unorganized and ineffective. Unlike ventricular fibrillation, which is lethal in minutes if not defibrillated, a person can live with atrial fibrillation, but the heart won’t pump efficiently causing fatigue, the backup of fluid into the lungs, and an increased risk of stroke.
- Electrical Mapping. The first part of the clinical procedure is to map the arrhythmia. Multiple electrodes on the catheter record the electrocardiogram throughout the atrium, locating the reentrant pathway or the focus (an isolated spot that initiates a wave front). If the arrhythmia is intermittent, then it may need to be triggered by electrical stimulation in order to map it.
- Ablation Sites. Once the arrhythmia is mapped, the doctor can determine where to ablate the tissue. Usually many isolated spots will be ablated to create a large lesion, often located around the pulmonary veins where many reentrant pathways occur.
 |
| My copy of Cardiac Electrophysiology: From Cell to Bedside, alongside IPMB. |
During RF catheter ablation, RF current passes through the tissue in close contact with the electrode and is resistively heated. The temperature of the tissue at the border of the lesion is reproducible in the 50°C to 55°C range. It is likely that the dominant model of myocardial injury is thermal, although electrical fields have been demonstrated to stun and kill cells depending on the field intensity. On inspection of the myocardial lesions, the tissue shows evidence of desiccation, inflammation, and microvascular injury, which certainly leads to ischemia. Late injury or recovery of the tissue at the lesion border zone may occur as a result of progression or resolution of inflammatory response or endothelial injury. On the cellular level, many possible mechanisms of myocyte damage exist, but membrane injury probably dominates. This may lead to cellular depolarization, intracellular Ca2+ overload, and cell death. Further damage to the cytoskeleton, cellular metabolism, and nucleus may occur at lower temperatures with more prolonged hyperthermia exposure. RF catheter ablation has been proven to be an effective clinical modality for the treatment of arrhythmias, but many of the basic pathophysiologic effects of this empirical procedure on the tissue and cellular level remain to be determined.
An interview with Dr. David Haines to discuss radiofrequency ablation.