|
| A figure from Barry et al., PNAS, 113:14133-14138, 2016. |
Barry et al.’s abstract states:
Magnetic fields from neuronal action potentials (APs) pass largely unperturbed through biological tissue, allowing magnetic measurements of AP dynamics to be performed extracellularly or even outside intact organisms. To date, however, magnetic techniques for sensing neuronal activity have either operated at the macroscale with coarse spatial and/or temporal resolution—e.g., magnetic resonance imaging methods and magnetoencephalography—or been restricted to biophysics studies of excised neurons probed with cryogenic or bulky detectors that do not provide single-neuron spatial resolution and are not scalable to functional networks or intact organisms. Here, we show that AP magnetic sensing can be realized with both single-neuron sensitivity and intact organism applicability using optically probed nitrogen-vacancy (NV) quantum defects in diamond, operated under ambient conditions and with the NV diamond sensor in close proximity (∼10 μm) to the biological sample. We demonstrate this method for excised single neurons from marine worm and squid, and then exterior to intact, optically opaque marine worms for extended periods and with no observed adverse effect on the animal. NV diamond magnetometry is noninvasive and label-free and does not cause photodamage. The method provides precise measurement of AP waveforms from individual neurons, as well as magnetic field correlates of the AP conduction velocity, and directly determines the AP propagation direction through the inherent sensitivity of NVs to the associated AP magnetic field vector.Here is my poor attempt to explain how their technique works; I must confess I don’t understand it completely. Nitrogen-vacancy defects in a diamond create a spin system that you can use for optically detected electron spin resonance. You shine light onto the system and detect the fluorescence with a photodiode. The shift in the magnetic resonance spectrum contained in the fluoresced light provides a measurement of the magnetic field. For our purposes we can think of the system as a black box that measures the magnetic field; new type of magnetometer.
Barry et al.’s paper started me thinking about the relative advantages and disadvantages of toroids versus optical methods for measuring magnetic fields of nerves.
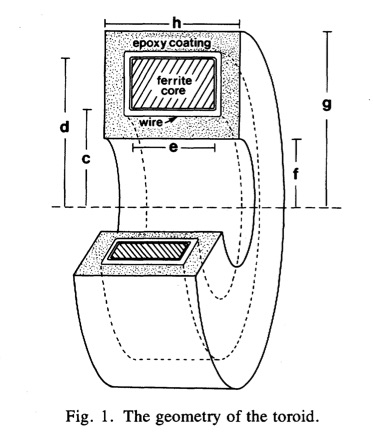
- A disadvantage of toroids is that you have to thread the nerve through the toroid center, which usually requires cutting the nerve. My PhD advisor John Wikswo created a “clip-on” toroid that avoids any cutting, but that technique never really caught on. In the optical method, you just drape the nerve over the detector like you would lie down on a bed. Winner: optical method.
- The toroid appears to provide a better signal-to-noise ratio than the optical method. Figure 3a in Roth and Wikswo (1985) shows that a signal of about 300 pT peak-to-peak can be detected with a signal-to-noise ratio of roughly 20 with no averaging (I don’t need to look at our 1985 paper to know this, I have a framed copy of this data on display in my office). Figure 2c of Barry et al. (2016) shows a 4000 pT peak-to-peak signal measured optically with a signal-to-noise ratio of about 10 after 600 averages. Perhaps this comparison is unfair because in 1985 we had spent several years optimizing our toroids, whereas this is a first measurement using the optical technique. Nevertheless, I think the toroids have a better signal-to-noise ratio. Winner: toroids.
- The optical technique appears to have better spatial resolution, although not dramatically so. Our toroids were typically one or two millimeters in size. In the optical method, the sensing layer is only 13 microns thick, but the length over which the detection occurs is two millimeters, so their method corresponds to a wide but small-diameter toroid. The spatial resolution of both methods could probably be improved, but once the size of the recording device is less than the length over which the action potential upstroke extends (about a millimeter) there is little to be gained by making the detector smaller. Both methods integrate the magnetic signal over the area of the device. Interestingly, the solution to last weeks new homework problem--integrating the biomagnetic field over the cross section of the toroid--is derived in the page 8 of the Barry et al.'s supplementary information. Winner: optical method.
- The optical method has better temporal resolution. Both, however, have temporal resolution adequate to record action potentials with an upstroke of a tenth of a millisecond or longer. The toroid does not record the time course of the magnetic field directly, but rather a mixture of the magnetic field and its derivative, and the technique requires a calibration pulse to get the “frequency compensation” correct. As best I can tell, the optical method measures the magnetic field directly. However, I believe any errors arising from frequency compensation of the toroids are small, and that the temporal resolution of both methods is fine. Winner: optical method.
- The toroids are encapsulated in epoxy, so they are biocompatible. I don’t know how biocompatible the optical method is, but I suspect it could be made biocompatible if a thin insulating layer covered the detecting layer, with a very small and probably negligible reduction in spatial resolution. Winner: tie.
- The question of convenience is tricky. I am accustomed to using the toroids, so that to me they are not difficult to use, whereas I have never tried the optical method. Nevertheless, I think toroids are more convenient. The optical method requires a static bias magnetic field, and toroids do not. Toroid measurements are insensitive to the exact position of the nerve within the toroid, whereas the optical method appears to be sensitive to the exact placement of the nerve on or near the detector. The optical method requires a spectroscopic analysis of the light signal; the toroid only needs an amplifier to record the current induced in the toroid winding. Finally, the optical method is based on magnetic splitting of spin states and magnetic resonance, whereas toroids rely on good old Faraday induction—my kind of physics. Although I consider toroids more convenient, I would not want to defend that opinion against a skeptical scientist holding a different view, because it is more a matter of taste than science. Winner: toroids.
Perhaps a more interesting comparison is for situations where you want to make two-dimensional measurements of current distributions. Barry et al. discuss how the optical method might be extended to do 2D imaging. The traditional method that would be the main competition would be Wikswo’s microSQUID or nanoSQUID: an array of small superconducting coils placed over a sample. But in that case you need cryogenics to keep the coils cold, and I could easily see how a room-temperature array of quantum defect detectors in diamond, recorded optically with a camera, might be simpler.
Both the optical method and toroids require placing a detector near the nerve, so both are invasive. However, if (IF!) the biomagnetic field can be used as the gradient field for magnetic resonance imaging (see my June 3 post) then that technique becomes totally noninvasive. Winner: MRI.