 |
| A word cloud based on Intermediate Physics for Medicine and Biology. |
I’ll let you decide if this word cloud is profound or silly. It was fun, and I like to share fun things with the readers of IPMB. Enjoy!
|
| A word cloud based on Intermediate Physics for Medicine and Biology. |
 |
| Cell Biology by the Numbers, by Ron Milo and Rob Phillips. |
One of the central missions of our book is to serve as an entry point that invites the reader to explore some of the key numbers of cell biology. We hope to attract readers of all kinds—from seasoned researchers, who simply want to find the best values for some number of interest, to beginning biology students, who want to supplement their introductory course materials. In the pages that follow, we provide a broad collection of vignettes, each of which focuses on quantities that help us think about sizes, concentrations, energies, rates, information content, and other key quantities that describe the living world.One part of the book that readers of Intermediate Physics for Medicine and Biology might find useful is their “rules of thumb.” I reproduce a few of them here
• 1 dalton (Da) = 1 g/mol ~ 1.6 × 10−24 g.The book consists of a series of vignettes, each phrased as a question. Here is an excerpt form one.
• 1 nM is about 1 molecule per bacterial volume [E. coli has a volume of about 1 μm3].
• 1 M is about one per 1 nm3.
• Under standard conditions, particles at a concentration of 1 M are ~ 1 nm apart.
• Water molecule volume ~ 0.03 nm3, (~0.3 nm)3.
• A small metabolite diffuses 1 nm in ~1 ns.
Which is bigger, mRNA or the protein it codes for?It’s obvious once someone explains it to you. Here is another that I liked.
The role of messenger RNA molecules (mRNAs), as epitomized in the central dogma, is one of fleeting messages for the creation of the main movers and shakers of the cell—namely, the proteins that drive cellular life. Words like these can conjure a mental picture in which an mRNA is thought of as a small blueprint for the creation of a much larger protein machine. In reality, the scales are exactly the opposite of what most people would guess. Nucleotides, the monomers making up an RNA molecule, have a mass of about 330 Da. This is about three times heavier that the average amino acid mass, which weighs in at ~110 Da. Moreover, since it takes three nucleotides to code for a single amino acid, this implies an extra factor of three in favor of mRNA such that the mRNA coding a given protein will be almost an order of magnitude heavier.
What is the pH of a cell?This one surprised me.
…Even though hydrogen ions appear to be ubiquitous in the exercise sections of textbooks, their actual abundance inside cells is extremely small. To see this, consider how many ions are in a bacterium or mitochondrion of volume 1 μm3 at pH 7. Using the rule of thumb that 1 nM corresponds to ~ 1 molecule per bacterial cell volume, and recognizing that pH 7 corresponds to a concentration of 10−7 M (or 100 nM), this means that there are about 100 hydrogen ions per bacterial cell…This should be contrasted with the fact that there are in excess of a million proteins in that same cellular volume.
What are the concentrations of free matabolites in cells?Somehow, I never realized how much glutamate is in cells. I also learned all sorts of interesting facts. For instance, a 5% by weight mixture of alcohol in water (roughly equivalent to beer) corresponds to a 1 M concentration. I guess the reason this does not wreak havoc on your osmotic balance is that alcohol easily crosses the cell membrane. Apparently yeast use the alcohol they produce to inhibit the growth of bacteria. This must be why John Snow found that during the 1854 London cholera epidemic, the guys working (and, apparently, drinking) in the brewery were immune.
…The molecular census of metabolites in E. coli reveals some overwhelmingly dominant molecular players. The amino acid glutamate wins out…at about 100 mM, which is higher than all other amino acids combined…Glutamate is negatively charged, as are most of the other abundant metabolites in the cell. This stockpile of negative charges is balanced mostly by a corresponding positively changed stockpile of free potassium ions, which have a typical concentration of roughly 200 mM.
…Say we drop a test substrate molecule into a cytoplasm with a volume equal to that of a bacterial cell. If everything is well mixed and there is no binding, how long will it take for the substrate molecule to collide with one specific protein in the cell? The rate of enzyme substrate collisions is dictated by the diffusion limit, which as shown above, is equal to ~ 109 s−1M−1 times the concentration. We make use of one of our tricks of the trade, which states that in E. coli, a single molecule (say, our substrate) has an effective concentration of about 1 nM (that is, 10−9 M). The rate of collisions is thus 109 s−1M−1 × 10−9 M. That is, they will meet within a second on average. This allows us to estimate that every substrate molecule collides with each and every protein in the cell on average about once per second.Each and every one, once per second! The beauty of this book, and the value of making these order-of-magnitude estimates, is to provide such insight. I cannot think of any book that has provided me with more insight than Cell Biology by the Numbers.
We leave our readers with the hope that they will find these and other questions inspiring and will set off on their own path to biological numeracy.
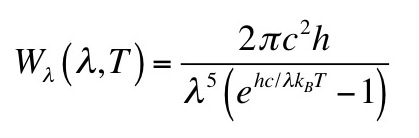
Problem 25 ½. Consider the limit of Planck’s law, Eq. 14.33, when hc/λ is much greater than kBT, an approximation known as the Wien exponential law.
(a) Derive the mathematical form of Wλ(λ,T) in this limit.
(b) Convert Wien’s law from a function of wavelength to a function of frequency, and determine the mathematical form of Wν(ν,T).
(c) Integrate Wν(ν,T) over all frequencies to obtain the total power emitted per unit area. Compare this result with the Stefan-Boltzmann law (Eq. 14.34). Derive an expression for the Stefan-Boltzmann constant in terms of other fundamental constants.
(d) Determine the frequency νmax corresponding to the peak in Wν(ν,T). Compare νmax/T with the value obtained from Planck’s law.
 |
| Subtle is the Lord, by Abraham Pais. |
Meanwhile,proposals for the correct form of [Wλ(λ,T)] had begun to appear as early as the 1860s. All these guesses may be forgotten except one, Wien’s exponential law, proposed in 1896…And the rest, as they say, is history.
Experimental techniques had sufficiently advanced by then to put this formula to the test. This was done by Friedrich Paschen from Hannover, whose measurements (very good ones) were made in the near infrared, λ = 1-8 μm (and T = 400 -1600 K). He published his data in January 1897. His conclusion: “It would seem very difficult to find another function…that represents the data with as few constants.” For a brief period, it appeared that Wien’s law was the final answer. But then, in the year 1900, this conclusion turned out to be premature…
2.6 The Chemostat
The chemostat is used by bacteriologists to study the growth of bacteria (Hagen 2010). It allows the rapid growth of bacteria to be observed over a longer time scale. Consider a container of bacterial nutrient of volume V. It is well stirred and contains y bacteria with concentration C = y/V. Some of the nutrient solution is removed at rate Q and replaced by fresh nutrient. The bacteria in the solution are reproducing at rate b. The rate of change of y is

Therefore the growth rate is slowed to

and can be adjusted by varying Q.However, Russ and I didn’t write any new homework problems for this section. If a topic is worth discussing in the text, then it’s worth creating homework problems to reinforce and extend that discussion. So, here are some new problems about the chemostat.
Problem 21.1. Often a chemostat is operated in steady state.
(a) Determine the solution removal rate Q required for steady state, as a function of the bacteria reproduction rate b and the container volume V, using Eq. 2.22. Determine the units of b, Q, and V, and verify that your expression for Q has the correct dimensions.
(b) If the rate Q is larger than the steady-state value, what is happening physically?
(c) Sometimes b varies with some external parameter (for example, temperature or glucose concentration), and you want to determine b as a function of that parameter. Suppose you can control Q and you can measure the number of bacteria y. Qualitatively design a way to determine b as your external parameter changes, assuming that for each value of the parameter your chemostat reaches steady state. (If unsure how to begin, take a look at Sec. 6.13.1 about the voltage clamp used in electrophysiology.)
Problem 21.2. Consider an experiment using a chemostat in which the bacteria's reproduction rate b slows as the number of bacteria y increases.
(a) Modify Eq. 2.22 so that “b” becomes “b (1 − y/y∞),” analogous to the logistic model (Sec. 2.10).
(b) Determine the value of y once the chemostat reaches steady state, as a function of Q, V, b, and y∞.
(c) Suppose your chemostat has a volume of 1.7 liters. You measure the steady state value of y (arbitrary units) for different values of Q (liters per hour), as shown in the table below. Plot y versus Q, and determine b and y∞.
Q y 0.2 11.64 0.4 9.47 0.6 7.31 0.8 5.14 1.0 2.98
Problem 21.3. Let the growth rate of the bacteria in your chemostat be limited by a small, constant amount of some essential metabolite, so the term “by” in Eq. 2.22 is replaced by a constant “a.”
(a) Find an expression for the solution removal rate Q in terms of a, the number of bacteria y, and the chemostat volume V, when the chemostat is in steady state.
(b) Determine the time constant governing how quickly the chemostat reaches steady state (Hint: see Sec. 2.8).
 |
| “Exponential Growth of Bacteria: Constant Multiplication Through Division,” by Stephen Hagen. |
Because the growth rate of the cell determines its size and chemical composition, a device that allows us to fine tune the growth rate will select the physiological properties of the cells. The bacterial chemostat is such a machine. In the chemostat a bacterial culture grows in a well-stirred vessel while a supply of fresh growth medium is fed into the vessel at a fixed flow rate Q (volume/time). At the same time, the medium (containing bacteria) is continuously withdrawn from the vessel at the same rate so as to maintain constant volume V. Thus, the bacterial population is continuously diluted at a rate D = Q/V. If this dilution rate exceeds the growth rate k [our b], the population is diluted, which allows its growth to accelerate until it matches the dilution rate, k = Q/V. (If D is too large, the culture will be diluted away entirely.) Therefore, the chemostat allows the experimenter to select the growth rate by selecting Q. Because it harnesses an exponential growth process to produce a tunable, steady output, we might think of the chemostat as the microbiological analog of a nuclear fission reactor. Interestingly, the chemostat reactor was first described by the physicist Leo Szilard (with Aaron Novick), who also (with Enrico Fermi) patented the nuclear reactor."I like the analogy to the nuclear reactor. Adjusting the flow rate in a chemostat is like pulling the cadmium control rods in and out of an atomic pile (except it’s less dangerous).
Neeraj Kapoor
June 3 at 1:36pm
Yesterday, during a conference with Elon Musk at a coding conference, he mentioned something about Neural Lacing (this group at harvard seems to be one of the few major groups working on it...http://cml.harvard.edu/) . I'm wondering if you have any knowledge of this Brad Roth and if so, if you could do a blog post on it.After a bit of googling, I found a Newsweek article about neural lacing, Elon Musk, and the coding conference.
Billionaire polymath Elon Musk has warned that humans risk being treated like house pets by artificial intelligence (AI) unless they implant technology into their brains.So what does all this talk about neural lacing mean, and how does it relate to Intermediate Physics for Medicine and Biology? As best I can tell, neural lacing would be used to monitor and excite nerves. The technology to stimulate nerves already exists, and is described in Chapter 7 of IPMB.
Musk believes that a technology concept known as “neural lace” could act as a wireless brain-computer interface capable of augmenting natural intelligence.
Speaking at the Code Conference in California on Wednesday, Musk said a neural lace could work “well and symbiotically” with the rest of a human’s body.
“I don’t love the idea of being a house cat, but what’s the solution? I think one of the solutions that seems maybe the best is to add an AI layer,” Musk said.
The information that has been developed in this chapter can also be used to understand some of the features of stimulating electrodes. These may be used for electromyographic studies; for stimulating muscles to contract called functional electrical stimulation (Peckham and Knutson 2005); for a cochlear implant to partially restore hearing (Zeng et al. 2008); deep brain stimulation for Parkinson’s disease (Perlmutter and Mink 2006); for cardiac pacing (Moses and Mullin 2007); and even for defibrillation (Dosdall et al. 2009). The electrodes may be inserted in cells, placed in or on a muscle, or placed on the skin.The best example of what I think Mr. Musk is talking about is the cochlear implant. A microphone records sound and analyzes it with a computer, which decides what location along the auditory nerve it should stimulate in order to fool the brain into thinking the ear heard that sound. For this technique to work, electrode arrays must be implanted in the cochlea so different spots can be stimulated, mimicking the sensitivity of different locations along the cochlea to different frequencies of sound.
Seamless and minimally invasive three-dimensional interpenetration of electronics within artificial or natural structures could allow for continuous monitoring and manipulation of their properties. Flexible electronics provide a means for conforming electronics to non-planar surfaces, yet targeted delivery of flexible electronics to internal regions remains difficult. Here, we overcome this challenge by demonstrating the syringe injection (and subsequent unfolding) of sub-micrometre-thick, centimetre-scale macroporous mesh electronics through needles with a diameter as small as 100 μm. Our results show that electronic components can be injected into man-made and biological cavities, as well as dense gels and tissue, with [greater than] 90% device yield. We demonstrate several applications of syringe-injectable electronics as a general approach for interpenetrating flexible electronics with three-dimensional structures, including (1) monitoring internal mechanical strains in polymer cavities, (2) tight integration and low chronic immunoreactivity with several distinct regions of the brain, and (3) in vivo multiplexed neural recording. Moreover, syringe injection enables the delivery of flexible electronics through a rigid shell, the delivery of large-volume flexible electronics that can fill internal cavities, and co-injection of electronics with other materials into host structures, opening up unique applications for flexible electronics.Is neural lacing science or science fiction? Hard to say. I am skeptical that in the future we will all have electrode arrays hardwired into our brains. But 50 years ago I would have been skeptical that cochlear implants could restore hearing to the deaf. I will reserve judgment, except to say that if neural lacing is developed, I am certain it will be based on the basic concepts Russ Hobbie and I discuss in Intermediate Physics for Medicine and Biology. That is the beauty of the book: it teaches the fundamental principles upon which you can build the amazing biomedical technologies of the future.
Syllabus, Biological PhysicsFall 2015
Class: Physics 325, MWF, 8:00–9:07, 378 MSC
Instructor: Brad Roth, Dept. Physics, 166 Hannah Hall, 370-4871, roth@oakland.edu, fax: 370-3408, office hours MWF, 9:15–10:00, https://files.oakland.edu/users/roth/web
Text: Intermediate Physics for Medicine and Biology, 5th Edition, by Hobbie and Roth (An electronic version of this book is available for free through the OU library)
Book Website: https://files.oakland.edu/users/roth/web/hobbie.htm (get the errata!).
Book Blog: http://hobbieroth.blogspot.com
Goal: To understand how physics influences and constrains biology
Grades
Point/Counterpoint 5 % Exam 1 Feb 5 20 % Chapters 1–3 Exam 2 March 18 20 % Chapters 4–6 Exam 3 April 20 20 % Chapter 7, 8, 10 Final Exam April 20 10 % Comprehensive Homework 25 %
Schedule
Sept 4 Introduction Sept 9, 11 Chapter 1 Mechanics, Fluid Dynamics Sept 14–18 Chapter 2 Exponential, Scaling Sept 21–25 Chapter 3 Thermodynamics Sept 28–Oct 2 Exam 1 Oct 5–9 Chapter 4 Diffusion Oct 12–16 Chapter 5 Osmotic Pressure Oct 19–23 Chapter 6 Electricity and Nerves Oct 26–30 Exam 2 Nov 2–6 Chapter 7 Extracellular Potentials Nov 9–13 Chapter 8 Biomagnetism Nov 16–20 Chapter 10 Heart Arrhythmias, Chaos Nov 23, 25 Chapter 10 Feedback Nov 30–Dec 4 Chapter 10 Feedback Dec 7 Review Dec 9 Final Exam
Homework
Chapter 1: 6, 7, 8, 16, 17, 33, 40, 42 due Wed, Sept 16 Chapter 2: 3, 5, 10, 29, 42, 46, 47, 48 due Wed, Sept 23 Chapter 3: 29, 30, 32, 33, 34, 40, 47, 48 due Wed, Sept 30 Chapter 4: 7, 8, 12, 20, 22, 23, 24, 41 due Wed, Oct 14 Chapter 5: 1, 3, 5, 6, 7, 8, 10, 16 due Wed, Oct 21 Chapter 6: 1, 2, 22, 28, 37, 41, 43, 61 due Wed, Oct 28 Chapter 7: 1, 10, 15, 24, 25, 36, 42, 47 due Wed, Nov 11 Chapter 8: 3, 10, 24, 25, 27, 28, 29, 32 due Wed, Nov 18 Chapter 10: 12, 16, 17, 18, 40, 41, 42, 43 due Wed, Dec 2
Syllabus, Medical PhysicsWinter 2016
Class: Physics 326, MWF, 10:40–11:47, 204 DH
Instructor: Brad Roth, Department of Physics, 166 HHS, (248) 370-4871, roth@oakland.edu, fax: (248) 370-3408, office hours MWF 9:30–10:30, https://files.oakland.edu/users/roth/web.
Text: Intermediate Physics for Medicine and Biology, 5th Edition, by Hobbie and Roth. An electronic version of the textbook is available through the OU library.
Book Website: https://files.oakland.edu/users/roth/web/hobbie.htm (get the errata!).
Book Blog: http://hobbieroth.blogspot.com
Goal: To understand how physics contributes to medicine
Grades
Point/Counterpoint 5 % Exam 1 Feb 5 20 % Chapters 13–15 Exam 2 March 18 20 % Chapters 16, 11–12 Exam 3 April 20 20 % Chapter 17, 18 Final Exam April 20 10 % Homework 25 %
Schedule
Jan 6, 8 Introduction Jan 11, 13, 15 Chpt 13 Sound and Ultrasound Jan 20, 22 Chpt 14 Atoms and Light Jan 25, 27, 29 Chpt 15 Interaction of Photons and Matter Feb 1, 3, 5 Exam 1 Feb 8, 10, 12 Chpt 16 Medical Uses of X rays Feb 15, 17, 19 Chpt 11 Least Squares and Signal Analysis Feb 22, 24, 26 Winter Recess Feb 29, March 2, 4 Chpt 12 Images March 7, 9, 11 Chpt 12 Images March 14, 16, 18 Exam 2 March 21, 23, 25 Chpt 17 Nuclear Medicine March 28, 30, Apr 1 Chpt 17 Nuclear Medicine April 4, 6, 8 Chpt 18 Magnetic Resonance Imaging April 11, 13, 15 Chpt 18 Magnetic Resonance Imaging April 18 Conclusion April 20 Final Exam
Homework
Chapter 13: 7, 10, 12, 21, 22, 27, 30, 36 due Fri, Jan 22 Chapter 14: 4, 5, 16, 21, 22, 47, 48, 49 due Wed, Jan 27 Chapter 15: 2, 4, 5, 10, 12, 14, 15, 16 due Wed, Feb 3 Chapter 16: 4, 5, 7, 16, 19, 20, 22, 31 due Wed, Feb 17 Chapter 11: 9, 11, 15, 20, 21, 36, 37, 41 due Wed, Mar 2 Chapter 12: 7, 9, 10, 23 due Wed, Mar 9 Chapter 12: 25, 32, 34, 35, and 27 (extra credit) due Wed, Mar 16 Chapter 17: 1, 2, 7, 9, 14, 17, 20, 22 due Wed, Mar 30 Chapter 17: 29, 30, 40, 54, 57, 58, 59, 60 due Wed, Apr 6 Chapter 18: 9, 10, 13, 14, 15, 18, 35, 49 due Wed, Apr 13
Point/Counterpoint articles
Jan 8: The 2014 initiative is not only unnecessary but it constitutes a threat to the future of medical physics. Med Phys, 38:5267–5269, 2011.
Jan 15: Ultrasonography is soon likely to become a viable alternative to x-ray mammography for breast cancer screening. Med Phys, 37:4526–4529, 2010.
Jan 22: High intensity focused ultrasound may be superior to radiation therapy for the treatment of early stage prostate cancer. Med Phys, 38:3909–3912, 2011.
Jan 29: The more important heavy charged particle radiotherapy of the future is more likely to be with heavy ions rather than protons. Med Phys, 40:090601, 2013.
Feb 12: The disadvantages of a multileaf collimator for proton radiotherapy outweigh its advantages. Med Phys, 41:020601, 2014.
Feb 19: Low-dose radiation is beneficial, not harmful. Med Phys, 41:070601, 2014.
March 4: Recent data show that mammographic screening of asymptomatic women is effective and essential. Med Phys, 39:4047–4050, 2012.
March 11: PDT is better than alternative therapies such as brachytherapy, electron beams, or low-energy x rays for the treatment of skin cancers. Med Phys, 38:1133–1135, 2011.
March 25: Submillimeter accuracy in radiosurgery is not possible. Med Phys, 40:050601, 2013.
April 1: Within the next ten years treatment planning will become fully automated without the need for human intervention. Med Phys, 41:120601, 2014.
April 8: Medical use of all high activity sources should be eliminated for security concerns. Med Phys, 42:6773, 2015.
April 15: MRI/CT is the future of radiotherapy treatment planning. Med Phys, 41:110601, 2014.
Much recent research has focused on using MRI to image neural activity directly, rather than through changes in blood flow (Bandettini et al. 2005). Two methods have been proposed to do this. In one, the biomagnetic field produced by neural activity (Chap. 8) acts as the contrast agent, perturbing the magnetic resonance signal. Images with and without the biomagnetic field present provide information about the distribution of neural action currents. In an alternative method, the Lorentz force (Eq. 8.2) acting on the action currents in the presence of a magnetic field causes the nerve to move slightly. If a magnetic field gradient is also present, the nerve may move into a region having a different Larmor frequency. Again, images taken with and without the action currents present provide information about neural activity. Unfortunately, both the biomagnetic field and the displacement caused by the Lorentz force are tiny, and neither of these methods has yet proved useful for neural imaging. However, if these methods could be developed, they would provide information about brain activity similar to that from the magnetoencephalogram, but without requiring the solution of an ill-posed inverse problem that makes the MEG so difficult to interpret.
 |
| “Direct Neural Current Imaging in an Intact Cerebellum with Magnetic Resonance Imaging,” by Sundaram et al. |
Functional study of the human brain has become possible with advances in non-invasive neuroimaging methods. The most widely used technique is blood oxygenation level-dependent functional MRI (BOLD-fMRI). Although BOLD-fMRI is a powerful tool for human brain activity mapping, it does not measure neuronal signals directly. Rather, it images slow local hemodynamic changes correlated with neuronal activity through a complex neurovascular coupling. At present, only electroencephalography (EEG) and magnetoencephalography (MEG) detect signals directly related to neuronal currents with a millisecond resolution. However, they estimate neuronal current sources from electrical potentials on the scalp or from magnetic fields outside the head, respectively. Measurement of these signals outside the brain leads to relatively poor spatial resolution due to ambiguity in inverse source estimation.After presenting their methods and results, Sundaram et al. write:
Our understanding of human brain function would significantly accelerate if it were possible to noninvasively detect neuronal currents inside the brain with superior spatiotemporal resolution. This possibility has led researchers to look for a method to detect neuronal currents with MRI. Many MRI approaches have been explored in the literature. Of these, the mechanism most commonly used is based on local changes in MR phase caused by neuronal magnetic fields. Electrical currents in active neurons produce magnetic fields (ΔB) locally within the tissue. The component of this field (ΔBz) along the main field (Bo) of the MR scanner alters the precession frequency of local water protons. This leads to a phase shift ΔΦ of the MR signal. For a gradient-echo (GE) sequence,
ΔΦ = γΔBzTE
where γ is the gyromagnetic ratio for hydrogen (2π × 42.58 MHz/T for protons) and TE is the echo time. According to Biot-Savart's law, ΔBz(t) is proportional to the current density J(t) produced by a population of neurons in the local region of the tissue. Thus, measurements of the phase shift ΔΦ can be used to directly estimate neuronal currents in the brain.
Many attempts have been made to detect neuronal currents in human subjects in vivo, but the results so far are inconclusive. The literature contains several reports of positive results which conflict with reports of negative results. This difficulty is presumably due to confounding factors such as blood flow, respiration and motion. Theoretical models, phantoms and cell culture studies indicate that it should be possible to detect neuronal currents with MRI in the absence of physiological noise sources.
Although these studies indicate that MRI technology should have enough sensitivity to detect neural currents, two types of key evidence are still lacking for demonstrating how MRI can be useful for neural current imaging: (1) there are no data showing that the phase shift is timelocked to some measure of population activity and that the phase shift time course matches that of a concurrently recorded local field potential (LFP), and (2) there is still no report showing how the phase shift data can be used to estimate the neuronal current distribution in the brain tissue, even though this should be the goal for neural current imaging.
Our work demonstrates that it is possible to measure an MR phase shift time course matching that of the simultaneously recorded evoked LFP in an isolated, intact whole cerebellum of turtle, free of physiological noise sources. We show how these MR phase maps can be used to estimate the neuronal current distribution in the active region in the tissue. We show that this estimated current distribution matches the distribution predicted based on spatial LFP maps. We discuss how these results can provide an empirical anchor for future development of techniques for in vivo neural current imaging.
We demonstrated that the ΔΦ can be detected reliably in individual cerebelli and that this phase shift is time-locked to the concurrently recorded LFP. The temporal waveform of the ΔΦ matched that of the LFP. Both the MR signal and LFP were produced by neuronal currents mediated by mGluRs. The measured values of ΔΦ in the individual time traces corresponded to local magnetic fields of 0.67–0.93 nT for TE = 26 ms. According to our forward solutions, these values correspond to a current dipole moment density q of 1–2 nA m/mm2,which agrees with the reported current density of 1–2 nA m/mm2 determined on the basis of MEG signals measured 2 cm above the cerebellum.They conclude
We also show that the MR phase data can be used to estimate the active neuronal tissue. This second step is important if MRI were to be used for imaging neuronal current distributions in the brain. We were able to use the minimum norm estimation technique developed in the field of MEG to estimate the current distribution in the cerebellum responsible for the measured phase shift. The peak values of ΔΦ in the phase map averaged across 7 animals were 0.15° and −0.10°, corresponding to peak ΔB values of +0.37 nT and −0.25 nT, respectively. The empirically obtained group-average ΔΦ of 0.12° and ΔB of 0.30 nT are close to the predicted values of 0.2° and 0.49 nT assuming q = 1 nA m/mm2. The slightly smaller group-average ΔΦ and ΔB may be due to variability in the spatial phase map and responses across animals.
Our results for metabotropic receptor mediated evoked neuronal activity in an isolated whole turtle cerebellum demonstrate that MRI can be used to detect neuronal currents with a time resolution of 100 ms, approximately ten times greater than for BOLD-fMRI, and with a sensitivity sufficiently high for near single-voxel detection. We have shown that it is possible to detect the MR phase shift with a time course matching that of the concurrently measured local field potential in the tissue. Furthermore, we showed how these MR phase data can be used to accurately estimate the spatial distribution of the current dipole moment density in the tissue.I’ve been interested in this topic for a while, publishing on the subject with Ranjith Wijesinghe of Ball State University (2009, 2012) and Peter Basser of the National Institutes of Health (2009, 2014). My graduate student Dan Xu (2012) examined the use of MRI to measure electrical activity in the heart, where the biomagnetic fields are largest. I remain skeptical that magnetic resonance imaging can record neural activity of the human brain in a way as accurate as functional MRI using BOLD. Yet, this is the first claim to have measured the magnetic field of neurons using MRI that I believe. It’s a beautiful result and a landmark study. I hope that I’m wrong and the method does have the potential for clinical functional imaging.






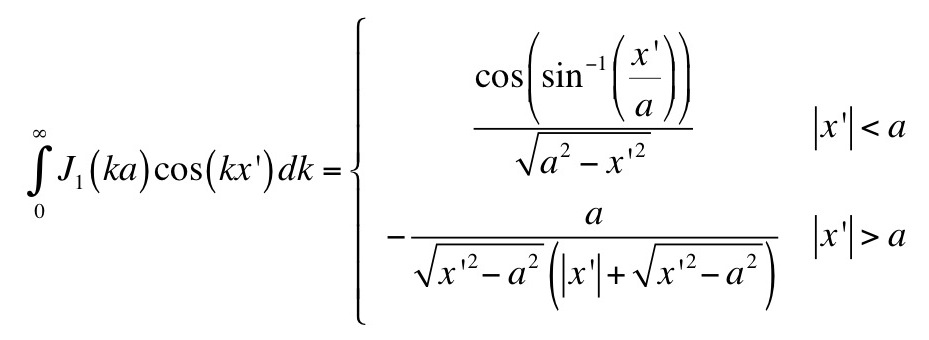

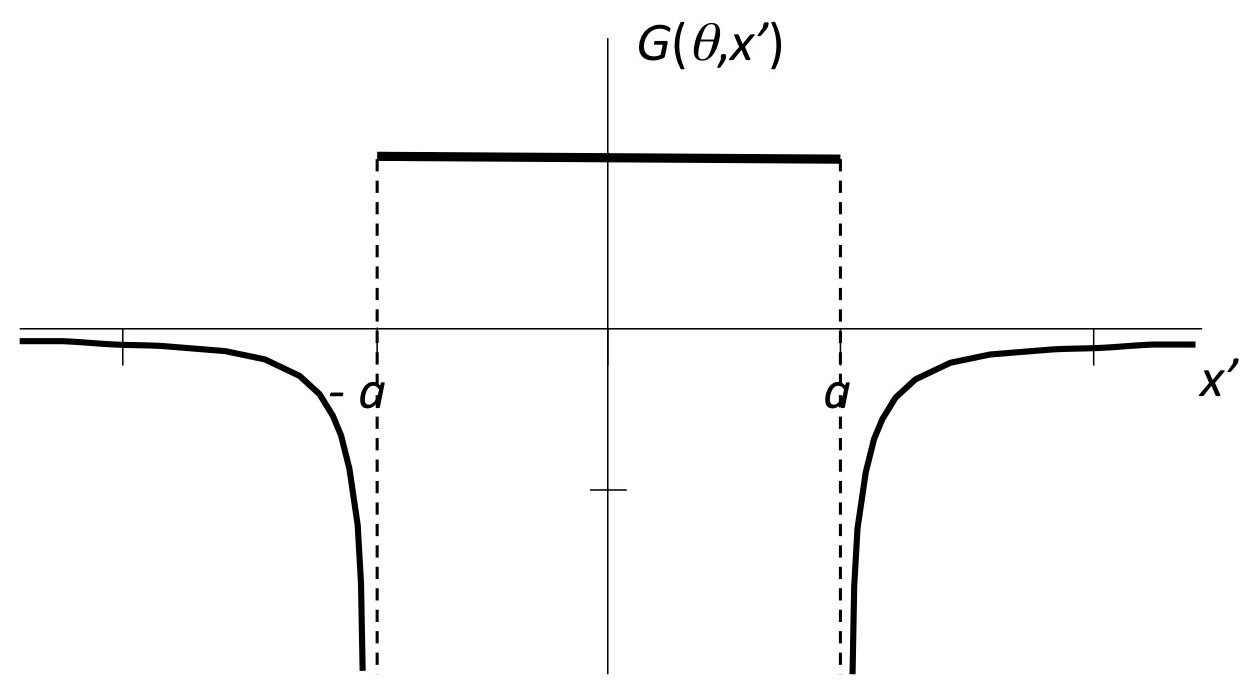






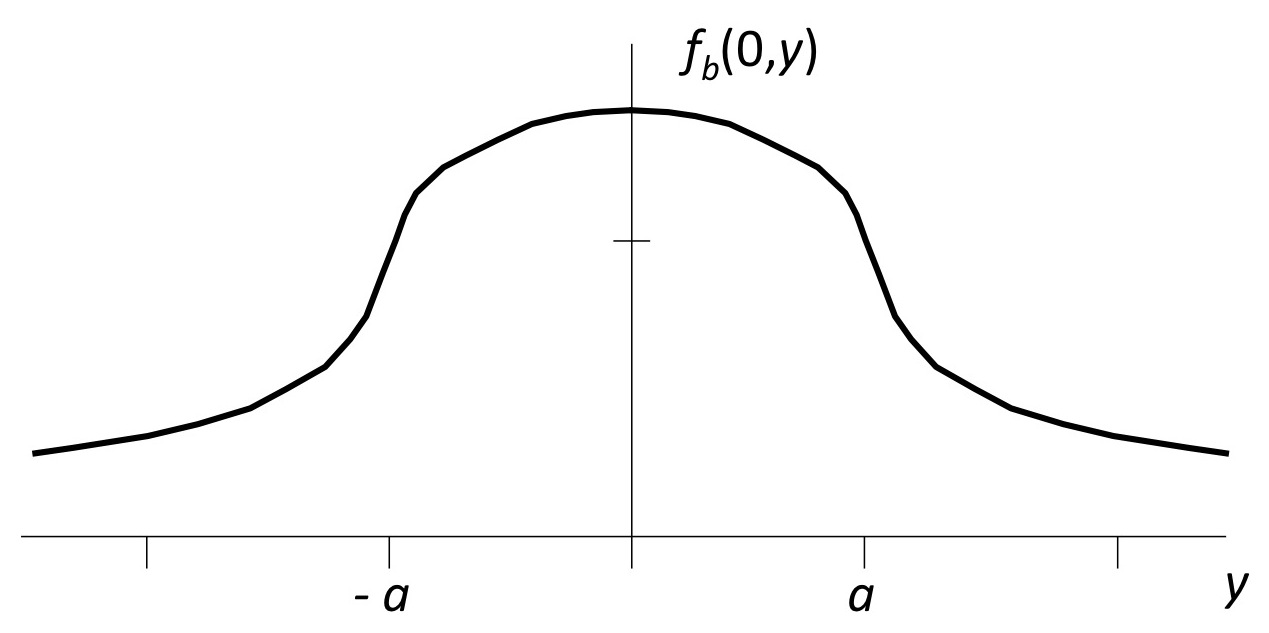
 |
| A five generation picture. |
 |
| All five editions of Intermediate Physics for Medicine and Biology. |
 |
| Suki with all five editions of Intermediate Physics for Medicine and Biology. |
 |
| All five editions of Intermediate Physics for Medicine and Biology. |
 |
| All five editions of Intermediate Physics for Medicine and Biology. |
 |
| Me holding all five editions of Intermediate Physics for Medicine and Biology. |
 |
| Trivial Pursuit. |
 |
| The game pieces for Trivial Pursuit. |
 |
| A game card for Trivial Pursuit. |
 |
| Trivial Pursuit. |


{kind=link}